きのこ驚きの秘密・その2 きのこの性 きのこの胞子を観察して、「タネ」としての働きのあることは1500年代に指摘されており、胞子には性的な役割のあることが推測されていました。1800年代までは、シダ属の植物のように「雄のきのこ」と「雌のきのこ」があると考えられていたようです。1788年にJ.ゲルトナーによって菌類には雌雄性がないと指摘されたのですが、なかなか信じてはもらえなかったようです。 その後、ビュリャールは子実体の中にシスチジアのあることを発見し、そのシスチジアには花粉と同様、精液が含まれ、胞子にシスチジアが精液を吹きかけることで受精すると考えたようです。1856年にシスチジアを徹底的に研究していたホフマンにより、ようやく「性」には無関係な器官であることが突き止められました。ちなみに、ホフマンはきのこの菌糸にクランプコネクション(かすがい連結)のあることを発見した人でもあります。しかし、ホフマンの主張は、当時、誰も信じてくれなかったようです。 1818年にエーレンベルクによりケカビの菌糸接合現象が発見され、2つの菌糸の「接合」が性現象(生殖)であることが発表されたことで、本格的にきのこの雌雄性の謎の解明に向けた研究に取りかかるきっかけとなったようです。これまで子実体にしか向けられなかった視線が、ようやく菌糸体に向けられるようになったのです。 1873年にリコンとローツェは、ヒトヨタケ属のきのこの菌糸体で菌糸融合が起きることを確認したのですが、当時の段階ではきのこの「性の謎」を解明するまでには至らなかったのです。 1895年にはハーパーにより、菌類には単相世代(栄養菌糸体)と複相世代(子実体)という2つの世代のあることが発見され、有性世代と無性世代とが交代することが明らかにされました。 1904年には、ハーバード大学のブレイクスリによりヘテロタリズム(性的異質接合性)による性現象が発見され、「雌雄性の謎」がようやく解明されることとなったのです。 さらに、1918年にハンス・クニープは、不稔系統のコロニーの接触で子実体が形成されることを明らかにし、交配で生じた菌糸にはクランプコネクションが形成されることを発見したのです。その後、パパチアンやレイパーらの交配型因子の研究成果で、菌類の自殖を抑制する交配システムとして、きのこの子実体には遺伝的に異なる2~4タイプの担子胞子を持つことが明らかとなりました。担子菌きのこの交配型には、1対の不和合性因子で制御される「2極性」と2対の不和合性因子で制御される「4極性」の2つのパターン(子のう菌きのこは2極性のみ)があり、2極性では50%、4極性では25%の確率で自家交配が抑制され、子孫に遺伝子型の多様性を増大させる効果のあることが解明されたのです。なお1930年には彼の有名なブラーにより、単相(1核)の菌糸体が複相(2核)の菌糸体と交配(接合)するダイ・モン交配の発見に至っているのです。 きのこの「雌雄性の謎」の解明の歴史は以上の通りで、1次菌糸同士の交配により子実体を生じることが解明された訳ですが、それでは1次菌糸には動植物で見られるような「雌雄」の区別はあるのでしょうか? 結論的には、「性」の区別はありますが菌糸には動植物のような形態的識別が困難なため、固定した雌雄の区別はありません。つまり、雄になる時もあれば、雌になる場合もあるということです。すなわち、核を受け入れる側の菌糸が「雌」であり、核を供与する側の菌糸が「雄」となるのです。交配したプレートからの菌糸の分離部位により、雌雄関係が逆転(正逆交配)してしまうのです。それゆえ、きのこの正逆交配においては細胞質遺伝子が母系遺伝することから、交配の際には雌雄関係が分かるようにしっかりと記録しておく必要があるのです。 菌類の性現象が初めて確認されたエーレンベルクが 描いたケカビの菌糸接合図(1820年) 引用文献 「キノコ・カビの研究史」人が菌類を知るまで(2010年) 著者:G.C.エインズワース/小川真 訳 発行所:京都大学学術出版会 きのこ菌糸の交配図(正逆交配)交配における核の表記例、拡大図 ◆ きのこの驚きの秘密、目次ページへ戻る ◆
きのこの性
きのこの胞子を観察して、「タネ」としての働きのあることは1500年代に指摘されており、胞子には性的な役割のあることが推測されていました。1800年代までは、シダ属の植物のように「雄のきのこ」と「雌のきのこ」があると考えられていたようです。1788年にJ.ゲルトナーによって菌類には雌雄性がないと指摘されたのですが、なかなか信じてはもらえなかったようです。
その後、ビュリャールは子実体の中にシスチジアのあることを発見し、そのシスチジアには花粉と同様、精液が含まれ、胞子にシスチジアが精液を吹きかけることで受精すると考えたようです。1856年にシスチジアを徹底的に研究していたホフマンにより、ようやく「性」には無関係な器官であることが突き止められました。ちなみに、ホフマンはきのこの菌糸にクランプコネクション(かすがい連結)のあることを発見した人でもあります。しかし、ホフマンの主張は、当時、誰も信じてくれなかったようです。
1818年にエーレンベルクによりケカビの菌糸接合現象が発見され、2つの菌糸の「接合」が性現象(生殖)であることが発表されたことで、本格的にきのこの雌雄性の謎の解明に向けた研究に取りかかるきっかけとなったようです。これまで子実体にしか向けられなかった視線が、ようやく菌糸体に向けられるようになったのです。
1873年にリコンとローツェは、ヒトヨタケ属のきのこの菌糸体で菌糸融合が起きることを確認したのですが、当時の段階ではきのこの「性の謎」を解明するまでには至らなかったのです。
1895年にはハーパーにより、菌類には単相世代(栄養菌糸体)と複相世代(子実体)という2つの世代のあることが発見され、有性世代と無性世代とが交代することが明らかにされました。
1904年には、ハーバード大学のブレイクスリによりヘテロタリズム(性的異質接合性)による性現象が発見され、「雌雄性の謎」がようやく解明されることとなったのです。
さらに、1918年にハンス・クニープは、不稔系統のコロニーの接触で子実体が形成されることを明らかにし、交配で生じた菌糸にはクランプコネクションが形成されることを発見したのです。その後、パパチアンやレイパーらの交配型因子の研究成果で、菌類の自殖を抑制する交配システムとして、きのこの子実体には遺伝的に異なる2~4タイプの担子胞子を持つことが明らかとなりました。担子菌きのこの交配型には、1対の不和合性因子で制御される「2極性」と2対の不和合性因子で制御される「4極性」の2つのパターン(子のう菌きのこは2極性のみ)があり、2極性では50%、4極性では25%の確率で自家交配が抑制され、子孫に遺伝子型の多様性を増大させる効果のあることが解明されたのです。なお1930年には彼の有名なブラーにより、単相(1核)の菌糸体が複相(2核)の菌糸体と交配(接合)するダイ・モン交配の発見に至っているのです。
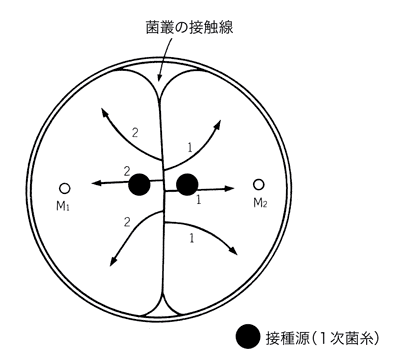
きのこの「雌雄性の謎」の解明の歴史は以上の通りで、1次菌糸同士の交配により子実体を生じることが解明された訳ですが、それでは1次菌糸には動植物で見られるような「雌雄」の区別はあるのでしょうか?
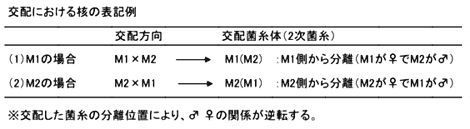
結論的には、「性」の区別はありますが菌糸には動植物のような形態的識別が困難なため、固定した雌雄の区別はありません。つまり、雄になる時もあれば、雌になる場合もあるということです。すなわち、核を受け入れる側の菌糸が「雌」であり、核を供与する側の菌糸が「雄」となるのです。交配したプレートからの菌糸の分離部位により、雌雄関係が逆転(正逆交配)してしまうのです。それゆえ、きのこの正逆交配においては細胞質遺伝子が母系遺伝することから、交配の際には雌雄関係が分かるようにしっかりと記録しておく必要があるのです。
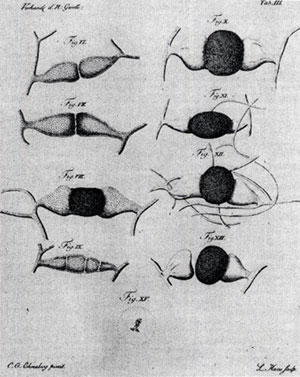
菌類の性現象が初めて確認されたエーレンベルクが
描いたケカビの菌糸接合図(1820年)
引用文献
「キノコ・カビの研究史」人が菌類を知るまで(2010年)
著者:G.C.エインズワース/小川真 訳
発行所:京都大学学術出版会
きのこ菌糸の交配図(正逆交配)
交配における核の表記例、拡大図